DESCRIÇÃO MORFOLÓGICA DOS RAMOS DO ARCO AÓRTICO E AORTA ABDOMINAL DO JAGUARUNDI (Herpailurus yagouaroundi)
MORPHOLOGICAL DESCRIPTION OF THE BRANCHES OF THE AORTIC ARCH AND ABDOMINAL AORTA OF THE JAGUARUNDI (Herpailurus yagouaroundi)
DOI: 10.24933/rep.v9i1.423
v. 9 n. 1 (2025)
SANTO, Iara Fernanda Lima do Espírito 1; FEIO, Júlia Vaz¹; BRANCO, Érika ²; LIMA, Ana Rita ²*
¹ Discente do Programa de Pós-graduação em Saúde e Produção Animal na Amazônia
(PPGSPAA- UFRA)- Belém, PA;
² Professora Doutora dos cursos de Medicina Veterinária e Zootecnia do Instituto da Saúde e
Produção Animal (ISPA) na Universidade Federal Rural da Amazônia (UFRA)- Belém, PA
* arlimavet@gmail.com
RESUMO. O jaguarundi (Herpailurus yagouaroundi) é um pequeno felino pertencente à classe Mammalia. Este trabalho tem como objetivo investigar e descrever os ramos colaterais do arco aórtico e da aorta abdominal. Foram estudados três espécimes de jaguarundi, nos quais o sistema arterial foi preenchido com solução de látex com contraste radiográfico e fixado em formaldeído a 10%. O arco aórtico de dois dos exemplares de jaguarundi analisados dividiu-se em dois ramos, o tronco braquiocefálico e a artéria subclávia esquerda. Em relação ao terceiro exemplar, foi encontrada configuração divergente, em que o arco aórtico originava o tronco braquiocefálico, com apresentação do tronco bicarotídeo e um tronco comum originando as duas artérias subclávias. Os ramos da aorta abdominal continham a artéria celíaca, que se trifurcava nas artérias hepática, gástrica esquerda e esplênica. Em seguida estava a artéria mesentérica cranial e, caudalmente a ela, as artérias renais, as artérias testiculares e a artéria mesentérica caudal. Por fim, estavam as artérias ilíacas externas e as artérias ilíacas internas e a artéria sacral mediana. Assim, conclui-se que as ramificações do arco ártico e da aorta abdominal de Jaguarundi apresentam semelhanças com as de cães e gatos, exceto pela presença de tronco bicarotídeo.
Palavras-chave: Artérias; Morfologia; Herpailurus yagouaroundi; Ramos colaterais; Tronco bicorotídeo.
ABSTRACT. The jaguarundi (Herpailurus yagouaroundi) is a small feline belonging to the class Mammalia. This work aims to investigate and describe the collateral branches of the aortic arch and abdominal aorta. Three specimens of jagouaroundi were studied, in which the arterial system was filled with latex solution with radiographic contrast and fixed in 10% formaldehyde. The aortic arch of two of the jaguarundi specimens analyzed divided into two branches, the brachiocephalic trunk and the left subclavian artery. Regarding the third specimen, a divergent configuration was found, in which the aortic arch originated the brachiocephalic trunk, with the presentation of the bicarotid trunk and a common trunk originating the two subclavian arteries. The branches of the abdominal aorta bore the celiac artery, which trifurcated into the hepatic, left gastric, and splenic arteries. Next was the cranial mesenteric artery and, caudally to it, the renal arteries, the testicular arteries and the caudal mesenteric artery. Finally, there were the external iliac arteries and the internal iliac arteries and median sacral artery. Thus, it is concluded that the ramifications of the arctic arch and the abdominal aorta of Jaguarundi have similarities to those in dogs and cats, except for the presence of a bicarotid trunk.
Keywords: Arteries; Morphology; Herpailurus yagouaroundi; Collateral branches; Bicarotid trunk.
INTRODUCTION
The jaguarundi (Herpailurus yagouaroundi) also known as red cat and gray cat is a small feline with an average weight of 5.2 kg, belonging to the Class Mammalia, Order Carnivora and Family Felidae (CASO et al., 2015). This species has an elongated body, with a uniform color, with brownish-black, grayish and yellowish red being the three basic types. It can also have a gray appearance (ADANIA et al., 2014).
H. yagouaroundi has a wide distribution throughout Brazil, albeit occurring at low population densities (ALMEIDA et al., 2013). It can have diurnal and nocturnal habits, with predominantly diurnal activity, and can be found in different biomes such as Cerrado, Caatinga, Pantanal and, in tropical and subtropical forests (ROCHA et al., 2017). The jaguarundi has a generalist diet, preying mainly on small mammals, especially small rodents, followed by birds and reptiles, mainly terrestrial species (TÓFOLI et al., 2009).
In Brazil, this species is not on the list of endangered animals, although its situation needs attention; the main threat being environmental degradation, which influences the exodus of these animals from their original habitat (CUBAS et al., 2014). Despite the wide distribution of the species, there are few studies about these wild cats (MICHALSKI et al., 2006), and most of them are related to macroscopic anatomy.
The Aortic Arch originates the Brachiocephalic Trunk, which is subdivided into arterial branches destined to irrigate the thoracic limbs, the regions of the head, the neck and the ventral part of the thorax; the abdominal aorta is a descending portion of the aorta, which passes from the thoracic cavity to the abdominal cavity through an opening in the diaphragm, called the aortic hiatus (KONIG; LIEBICH, 2021). It follows the roof of the abdomen, related to the caudal vena cava on the right and the psoas muscles on the left. Along its course, the abdominal aorta produces both visceral and parietal branches (DYCE et al., 2019).
Therefore, in view of the scarce literature available on morphological aspects of H. yagouaroundi and the importance of systemic anatomy for a physiological understanding of these animals, this work aims to systematically investigate and describe the collateral branches of the aortic arch and abdominal aorta in this species.
MATERIAL AND METHODS
Three adult male specimens of jaguarundi (Herpailurus yagouaroundi) killed by being run over were used, obtained from the Bauxite Mine - Paragominas-PA, under authorization SEMA-PA Nº 455/2009 and 522/2009. These were cryopreserved and sent to the Laboratory of Animal Morphological Research (Lama), from the Federal Rural University of the Amazon- UFRA.
First, the animals were thawed in running water for a period of approximately 4 hours. Next, the animals were dissected at the level of the aortic arch, through the 5th left intercostal space, which was cannulated and injected with Neoprene 650 latex solution, with red dye (Xadrez®) and radiographic contrast (barium sulfate) added to fill out the entire arterial vascular system. After this procedure, the specimens were fixed with a 10% formaldehyde aqueous solution through intramuscular, subcutaneous and intracavitary perfusion, keeping them submerged in the same fixative solution for a minimum period of seven days.
After the fixation process, an incision was made in the ventral plane of the animals, from the mentonian region to the pelvic region, followed by folding back the skin and muscles, which allowed access to the aortic arch and abdominal aorta and all its branches for identification and photographic recording of the arteries of interest in this study.
Finally, the animals were radiographed with an X-ray device (Intecal®, model CR-7), at a power of 100 kV (kilovoltage) and 100 mA (milliamperage); AGFA® brand 24 x 30 cm films were used, maintaining a distance between the focus and the film of 90 cm, with exposure of 50 KV, 100 mA and time 0.2 seconds, with the aim of helping to identify the collateral branches of the aortic arch and abdominal aorta.
RESULTS AND DISCUSSION
There are studies in the literature that describe the ramifications of the aortic arch in domestic and wild mammals. These comprise the main arteries that issue directly from the arch, with different types of vascular arrangements for each species. Basically, three models of ramifications are found. The first type corresponds only to the formation of the brachiocephalic trunk arising from the aortic arch and from it arise the right and left common carotid arteries and the right and left subclavian arteries, while the second arrangement is characterized by the appearance of two branches, the brachiocephalic trunk, followed by the left subclavian artery. Finally, in the third vascular model three collateral arteries of the aortic arch appear, the brachiocephalic trunk, the left common carotid artery and the left subclavian artery (DE OLIVEIRA et al., 2018).
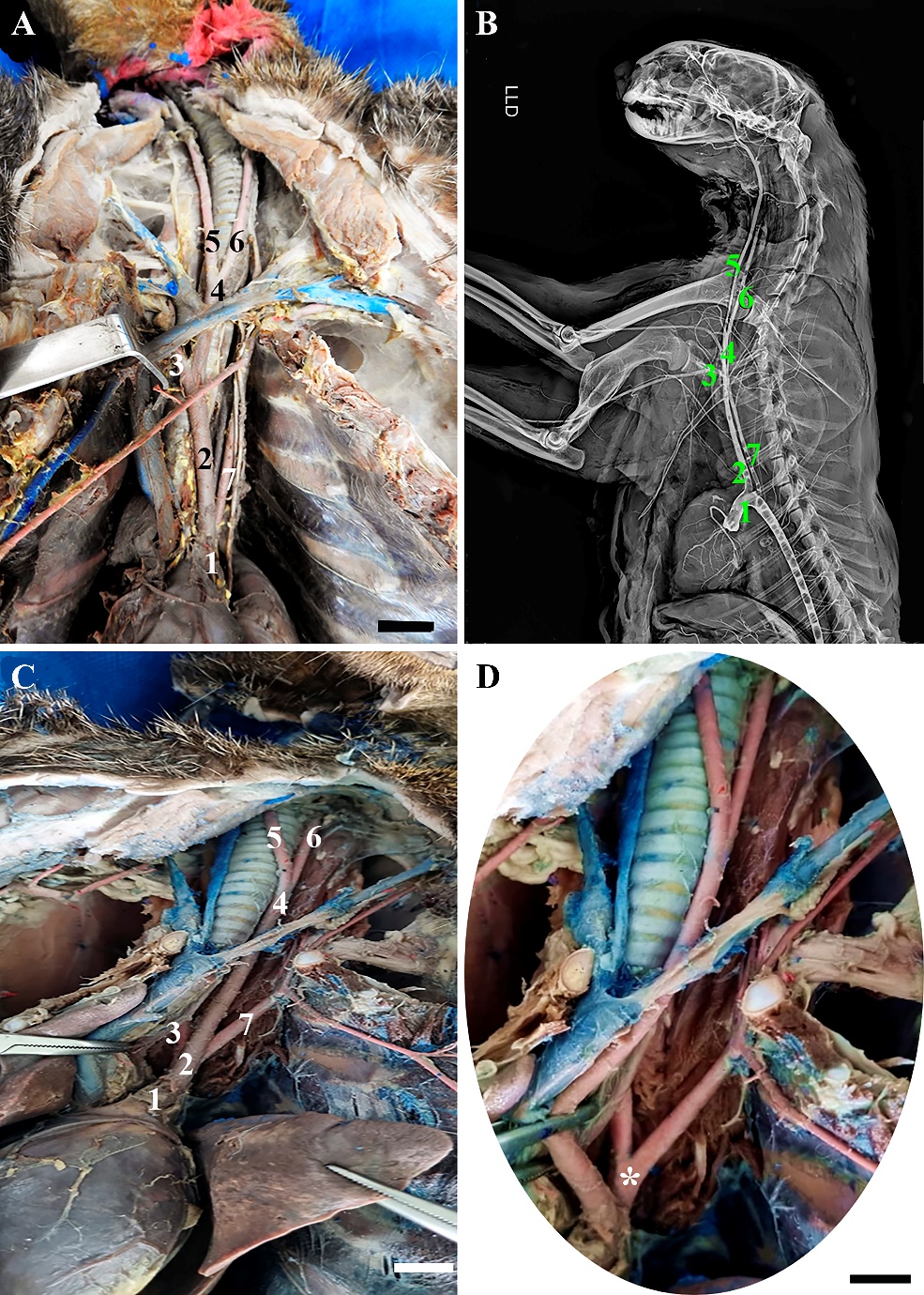
Among the H. yagouaroundi specimens analyzed, two specimens had two branches originating from the aortic arch (Table 1), located at the level of T5, the brachiocephalic trunk and the left subclavian artery. The brachiocephalic trunk first gave rise to the right subclavian artery and then the bicarotid trunk emitted at the level of the seventh cervical vertebra (C7), which in turn gave rise to the right and left common carotid arteries (Figure 1A). Similar to that described in dogs and cats, in which the arch originates from the brachiocephalic trunk and the left subclavian artery, while the brachiocephalic trunk emits the left common carotid arteries and right (DYCE et al., 2019), with the exception of the bicarotid trunk; other studies describe the configuration seen in H. yagouaroundi as the same as that found in the crab-eating raccoon (Procyon cancrivourus) (PEREIRA et al., 2016). The third specimen of H. yagouaroundi had a divergent configuration called the bissubclavian trunk in which the right and left subclavian arteries originated from a common trunk. This is like findings from studies carried out in dogs (VITUMS, 1962; GERRITY; MARTIN, 1980). In relation to the third specimen, it presented a divergent configuration, in which the aortic arch, also emitted at the level of the fifth thoracic vertebra, originated two branches, the branchiocephalic trunk, with the presence of the bicarotid trunk and the right and left common carotid arteries and a common trunk that bifurcated into the two right and left subclavian arteries (Figure 1C).
Table 1 – Description of the origin of the main branches of the aortic arch of the abdominal aorta of H. yagouaroundi.
|
Origin artery |
Vertebra of Origin |
|
|||
|
Brachiocephalic Trunk |
aortic arch |
T5 |
|
||
|
Left Subclavian Artery |
aortic arch |
T5 |
|
||
|
Bicarotid Trunk |
Brachiocephalic Trunk |
C7 |
|
||
|
Right Subclavian Artery * |
Brachiocephalic Trunk |
C7 |
|
||
|
Left common carotid artery |
bicarotid trunk |
C7 |
|
||
|
Right Common Carotid Artery |
bicarotid trunk |
C7 |
|
||
|
celiac artery |
Aorta Abdominal |
Between T13 e L1 |
|||
|
Left Gastric Artery |
|
|
|
||
|
hepatic artery |
celiac artery |
Between T13 e L1 |
|
||
|
Splenic artery |
|
|
|
||
|
Cranial Mesenteric Artery |
Aorta Abdominal |
Between T13 e L1 |
|
||
|
Renal arteries |
Aorta Abdominal |
L2 |
|
||
|
Caudal mesenteric artery |
Aorta Abdominal |
L6 |
|
||
|
Testicular arteries |
Aorta Abdominal |
L5 |
|
||
|
External Iliac Arteries |
|
|
|
||
|
Internal Iliac Arteries |
Aorta Abdominal |
L7 |
|
||
|
Median Sacral Artery |
|
|
|
||
*Only animal 3 presented the branching of the right subclavian artery parallel to the left subclavian artery originating from a common trunk.
The arrangement found in specimens of H. yagouaroundi showed a common trunk between the two carotids, as seen in lesser anteater (Tamandua tetradactyla) (PINHEIRO et al. 2012) and crab-eating fox (Cerdocyon thous) (LIMA et al., 2016), differing from the findings reported in ocelots (Leopardus pardalis) (MARTINS et al., 2010) and in dogs and cats, as there is no bicarotid trunk in these species (KONIG; LIEBICH, 2021). The aortic arch in wild dog specimens presented only two vessels, the brachiocephalic trunk and the left subclavian artery, from the first emerges the right subclavian artery and the bicarotid trunk, which will originate the left and right common carotid arteries (LIMA et al., 2016). Moreover, in some cavy specimens these same branches seen in H. yagouaroundi and crab-eating fox (Cerdocyon thous) were found, with the presence of the bicarotid trunk (OLIVEIRA et al., 2015).
In nutrias (Myocastor coypus), in most of the specimens surveyed by Campos et al. (2010) it was possible to visualize an aortic arch configuration similar to that found in this study, in which the aortic arch curved to the left, emitting the following branches: the brachiocephalic trunk and the left subclavian artery in 60% of the animals, in the remaining 40%, the sequence of the branches of the aortic arch was the brachiocephalic trunk, the left common carotid artery and the left subclavian artery, in relation to the brachiocephalic trunk.
Another study in which the arrangement of the branches of the divergent aortic arch of H. yagouaroundi can be observed is in the capuchin monkey (Sapajus apella), in which the aortic arch has a configuration like that found in some specimens of nutria, emitting as main branches the brachiocephalic trunk and the left common carotid and left subclavian arteries (FURTADO et al., 2017).
Figure 1– Photomacrographs showing the main branches of the aortic arch of H. yagouaroundi. In A), it is possible to observe the Aortic arch (1), Brachiocephalic trunk (2), Right subclavian artery (3), Bicarotid trunk (4), Right common carotid artery (5), Left common carotid artery (6) and Left subclavian artery (7). In B) right lateral projection radiographic image, note the same branches identified in figure A. In C) and D), we observe the main branches already described of the aortic arch of the third specimen, including the common trunk (*) that originates the two subclavias left and right (3 and 7). Scale bars: 1cm
The abdominal aorta of specimens of H. yagouaroundi emerged in the abdominal cavity after crossing the diaphragm in the region of the aortic hiatus. The first visceral branch emitted by the abdominal aorta was the celiac artery, located between the transition of the T13 and L1 vertebrae, which trifurcated, originating the hepatic, left gastric and splenic arteries that perform the vascularization of the liver, stomach and spleen, respectively. The hepatic artery was further divided into gastroduodenal and right gastric arteries. The cranial mesenteric artery was a unique and thicker branch that appeared soon afterwards in the caudal portion of the celiac artery, also emitted between the transition of the T13 and L1 vertebrae, and proceeding towards the mesentery, originating the caudal pancreaticoduodenal, jejunal and ileocolic arteries, which branched into the middle and left colic arteries (Figure 2).
As in most domestic animals (DYCE et al., 2019) and wild animals such as the capuchin monkey (Sapajus apella) (FURTADO et al., 2017), the celiac artery of H. yagouaroundi is the first branch of the abdominal aorta, which usually develops in three main branches, as described in carnivores, horses and ruminants (DYCE et al., 2019), as well as ocelots (Leopardus pardalis) (PINHEIRO et al. 2014) and squirrel monkeys (Saimiri sciureus) (SILVA et al., 2011). Although this arrangement of branches was not found in H. yagouaroundi, the left gastric and splenic arteries may arise from a common trunk, called the gastrosplenic trunk, as described in some specimens of domestic cats studied (XAVIER-SILVA et al., 2012).
Furthermore, the ramifications of the hepatic artery of H. yagouaroundi are like the configuration found in dogs and cats, in which it bifurcates into the right gastric and gastroduodenal arteries (DYCE et al., 2019). In the study by Xavier-Silva et al. (2013) carried out on 30 specimens of domestic cats, four animals had the origin of the celiac artery located between the 13th thoracic vertebra and the 1st lumbar vertebra, results like those found in H. yagouaroundi.
After the celiac trunk, the cranial mesenteric artery was identified in all animals, originating from the abdominal aorta and, caudally to the origin of the celiac trunk, being of greater caliber than this, corroborating the findings of Culau et al. (2008) in nutria and with Pinheiro et al. (2014) in ocelots. Towards the mesentery, the cranial mesenteric artery gives rise to the caudal pancreaticoduodenal arteries, jejunal arteries and ileocolic artery with a configuration similar to the studies by Santos et al (2004) in the Amazon tortoise (Podocnemis expansa), in the findings by Silva (2022) in bush dog (Cerdocyon thous) that presented the same ramifications and in the dog and cat (DYCE et al., 2019), and in the dog one can notice the branching of the ileocolic artery into middle colic and right colic arteries (BUDRAS et al., 2012) similar to that found in this study.
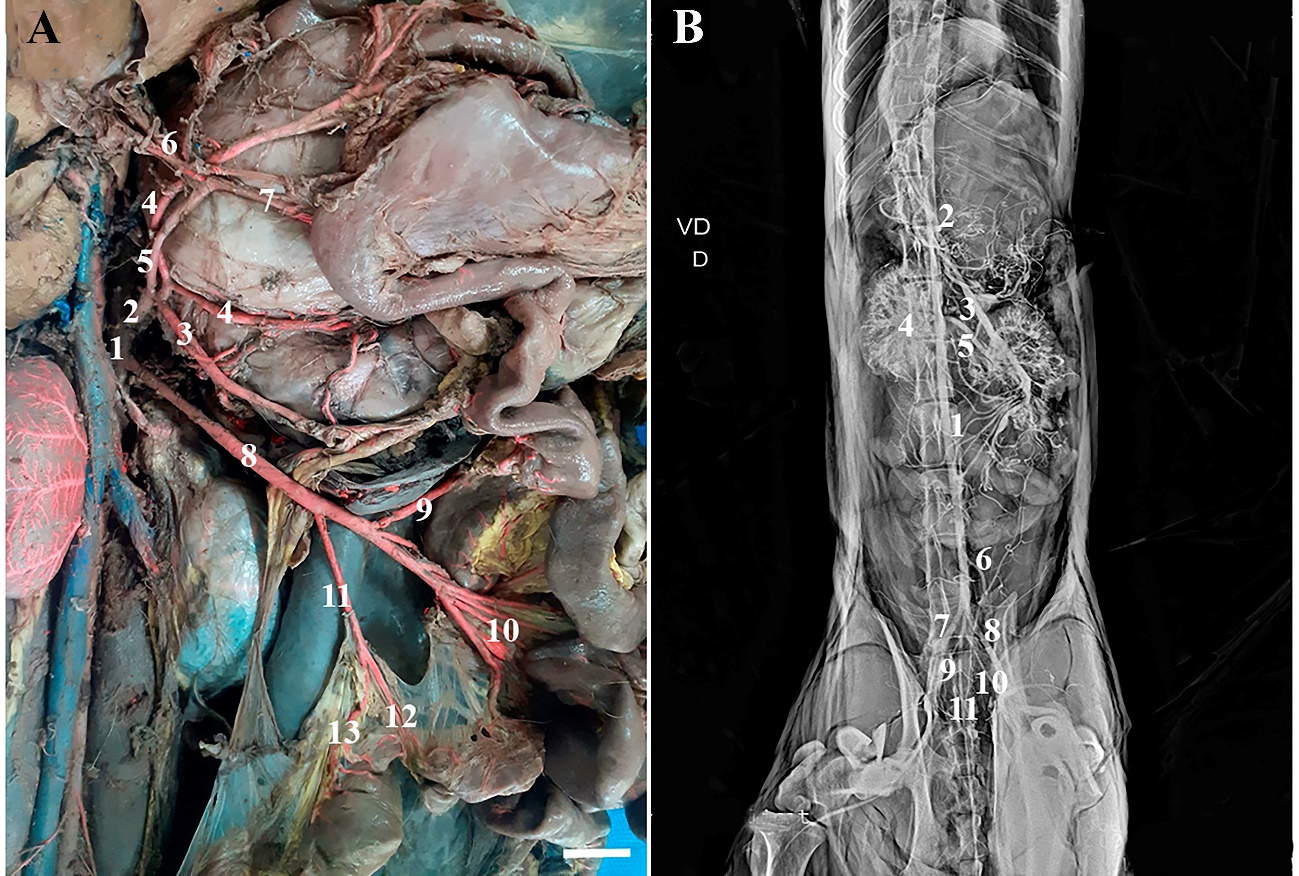 Figure 2 – Photomacrographs of the Abdominal Aorta and its collateral
branches in H. yagouaroundi. In A) Abdominal aorta (1),
Celiac artery (2), Splenic artery (3), Left gastric artery (4), Hepatic artery
(5), Gastroduodenal artery (6), A. right gastric (7), A. cranial mesenteric
(8), A. caudal pancreaticoduodenal (9), A. jejunal (10), A. ileocolic (11), A.
middle colic (12), A. left colic (13). In B) ventrodorsal
radiograph of branches of the abdominal aorta of a male H. yagouaroundi,
Abdominal Aorta (1), Celiac artery (2), Cranial mesenteric artery (3), Right
kidney (4), Left renal artery (5), Caudal mesenteric artery (6), Right external
iliac artery (7), Left external iliac artery (8), Right internal iliac artery
(9), Left internal iliac artery (10) and Median sacral artery (11). Scale bar:
1cm
Figure 2 – Photomacrographs of the Abdominal Aorta and its collateral
branches in H. yagouaroundi. In A) Abdominal aorta (1),
Celiac artery (2), Splenic artery (3), Left gastric artery (4), Hepatic artery
(5), Gastroduodenal artery (6), A. right gastric (7), A. cranial mesenteric
(8), A. caudal pancreaticoduodenal (9), A. jejunal (10), A. ileocolic (11), A.
middle colic (12), A. left colic (13). In B) ventrodorsal
radiograph of branches of the abdominal aorta of a male H. yagouaroundi,
Abdominal Aorta (1), Celiac artery (2), Cranial mesenteric artery (3), Right
kidney (4), Left renal artery (5), Caudal mesenteric artery (6), Right external
iliac artery (7), Left external iliac artery (8), Right internal iliac artery
(9), Left internal iliac artery (10) and Median sacral artery (11). Scale bar:
1cm
The right and left renal arteries appeared caudally to the cranial mesenteric artery, vessels that were large in caliber as found in the squirrel monkey (Saimiri sciureus) (SILVA et al., 2011), as well as in coati (BIIHRER et al., 2015). The renal arteries of H. yagouaroundi followed the pattern described in domestic animals, in which the right renal artery is slightly more cranial in relation to the left following the more cranial position of the right kidney, like dogs (GHOSHAL, 1986) and located at the level of the L2 vertebra (Figure 3A). This configuration it is also found in other wild animals, such as in studies with lesser anteaters (Tamandua tetradactyla) (MACEDO et al., 2013) and owl monkeys (Aotus azarae infulatus) (FARIA et al., 2016).
The left and right testicular arteries of H. yagouaroundi at the level of the L5 vertebra, were thin and long and arose ventrolaterally from the abdominal aorta (Figure 3A). The testicular arteries have a variable origin described in the literature. In most domestic animals, dogs, horses, pigs it is reported as a branch of the abdominal aorta (DYCE et al., 2019), similar to the findings in the present study, as well as in ocelots (L. pardalis) (PINHEIRO et al., 2014) that presented long and thin testicular arteries, originating from the abdominal aorta and different from that found in capuchin monkey (S. apella) (FURTADO et al., 2017) and lesser anteater (Tamandua tetradactyla) (MACEDO et al., 2013), which have an atypical configuration, in which the testicular arteries are collateral branches of the renal arteries. Furthermore, in studies with squirrel monkeys carried out by Silva et al. (2011), they describe another configuration, in which the testicular arteries in the species originate from the medial face of the internal iliac arteries and are short and thin.
In H. yagouaroundi, it was observed that the caudal mesenteric artery originated on the ventral side, close to the terminal portion of the abdominal aorta, dividing into the left colic artery and cranial rectal arteries (Figure 3B), as well as in dogs and cats (DYCE et al., 2019). This arrangement of origin and configuration pattern of the vessels can also be found in wild animals such as the Kerodon rupestris (QUEIROZ et al., 2011), wild boar (Sus scrofa scrofa) (GOMES et al., 2013), pacas (Cuniculus paca) (de Souza Marques et al., 2013), ocelot (Leopardus pardalis) (PINHEIRO et al., 2014), night monkey (Aotus azarae infulatus) (FARIA et al., 2016) and crab-eating fox (Cerdocyon thous) (SILVA, 2022).
As for the terminal portion of the abdominal aorta, some divergences are observed when compared to domestic carnivores (DYCE et al., 2019), since in these animals the caudal portion emits the external and internal iliac arteries all issued at the L7 level, in addition to the median sacral artery, in the specimens studied of the jaguarundi the external iliac arteries arise and then, through a common trunk originating from the abdominal aorta, the internal iliac arteries originate and, finally, the median sacral artery (Figure 3B), similar to that found in coatis (Nasua nasua) (FELIPE et al., 2017). This disposition differs from descriptions in nutria (Myocastor coypus) (CULAU et al., 2008), night monkey (Aotus azarae infulatus) (FARIA et al., 2016), capuchin monkey (Sapajus apella) (FURTADO et al., 2017) in which the internal iliac arteries arise from the branch of the external iliac arteries.
Regarding the location of the vertebrae of the main branches analyzed, studies are scarce, in specimens analyzed of sloths (Bradypus variegatus), the aortic arch originated at the level of the fourth thoracic vertebra (T4), close to that found in H. yagouaroundi, and the other branches originating from the aortic arch ranged from T1 to T3 in the studied specimens (ALBUQUERQUE et al., 2018).
The arrangement of branches of the abdominal aorta has been described in certain wild animals. In the study with Pampas foxes (Lycalopex gymnocercus) (LEÃO et al., 2017), the celiac artery in the female originated at the first lumbar vertebra (L1) and in the male at the level of the second (L2), findings similar to another study carried out on the same species in which 64% of the cases the celiac artery originated at the level of the second lumbar vertebra (L2), while the rest were between L1 and L2, or only at L1 (NETO et al., 2019).
According to Silva et al. (2011), the cranial mesenteric artery, found in the specimens of squirrel monkeys studied, was located at the level of L2, the right and left renal arteries, both were located close to L2 and L3, Similar to the arrangement of the renal arteries of H. yagouaroundi at the levels of the second lumbar vertebra (L2), the caudal mesenteric artery originated in L3 and the internal and external iliac arteries were inserted in L6, which differs from the results found in this study, in which Caudal mesenteric artery is located at L6 and the external and internal iliac arteries at L7. Additionally, the configuration found in cats by Silva et al. (2011) is like that described in night monkeys (Aotus azarae infulatus) by Faria et al. (2016), in which the caudal mesenteric arteries and external and internal iliac arteries were inserted at L3 and L6, respectively.
Although this study provided interesting results on the ramifications of the aortic arch and abdominal aorta in H. yagouaroundi, some limitations should be considered. Firstly, due to the specimens being victims of being run over and coming from mining areas, the study contained few specimens of the H. yagouaroundi species available to carry out the research, which may restrict the generalization of the results to the entire population of the species. Furthermore, describing the morphology of the main branches of the aortic arch and abdominal aorta of jaguarandi proved to be challenging, since most descriptions found in the literature are of domestic animals, in addition to the atypical anatomical configuration of the aortic arch observed in the third specimen of the species. Therefore, these limitations suggest the need for further research related to the subject.
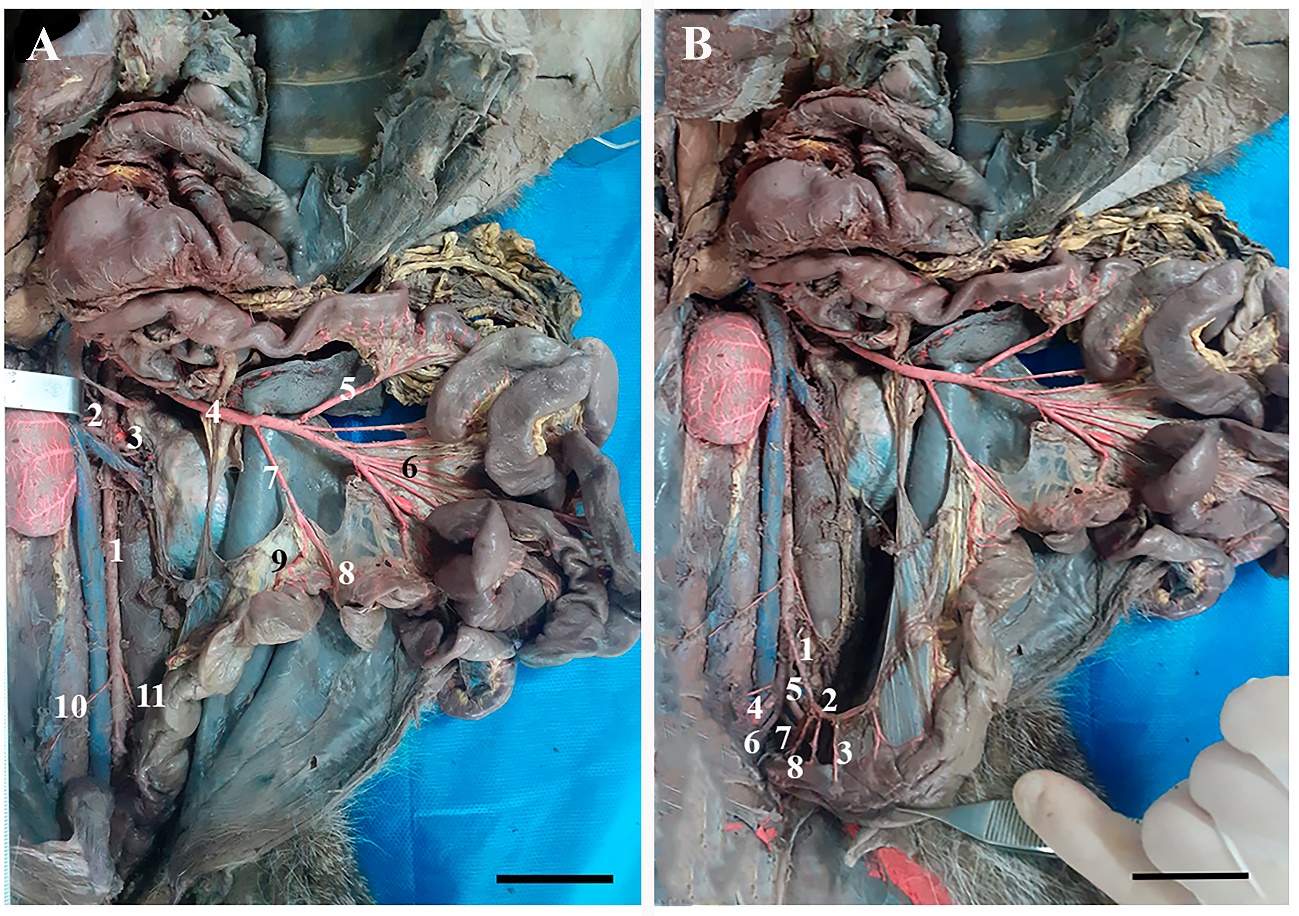 Figure 3 –
Photomacrographs
of the Abdominal Aorta and its collateral branches in H. yagouaroundi.
In A) the abdominal aorta (1), right renal artery (2), left renal artery
(3), cranial mesenteric artery (4), caudal pancreaticoduodenal artery (5),
jejunal arteries (6), A. ileocolic (7), A. middle colic (8), A. left colic (9),
A. right testicular (10), A. left testicular (11). In B) note the caudal
mesenteric arteries (1), left colic (2), cranial rectal (3), right external
iliac (4), left external iliac (5), right internal iliac (6), left internal
iliac (7) and median sacral (8). Scale bars: 3 cm.
Figure 3 –
Photomacrographs
of the Abdominal Aorta and its collateral branches in H. yagouaroundi.
In A) the abdominal aorta (1), right renal artery (2), left renal artery
(3), cranial mesenteric artery (4), caudal pancreaticoduodenal artery (5),
jejunal arteries (6), A. ileocolic (7), A. middle colic (8), A. left colic (9),
A. right testicular (10), A. left testicular (11). In B) note the caudal
mesenteric arteries (1), left colic (2), cranial rectal (3), right external
iliac (4), left external iliac (5), right internal iliac (6), left internal
iliac (7) and median sacral (8). Scale bars: 3 cm.
CONCLUSION
From the results observed, it was noted that many branches of the aortic arch and abdominal aorta of jaguarundi are like those found in dogs and cats, except for the presence of a bicarotid trunk that originated the left and right common carotid arteries in jaguarundi. Although they are not specifically linked to wild animals, these anatomical variations must be identified and described, since clinical and surgical practice, predominantly in surgical approaches to the abdominal cavity in these animals, has become more recurrent in veterinary clinics.
REFERENCES
ALBUQUERQUE, P.V.; SENA, D.G.F.; BRAZ, R.S.; MESQUITA, E.P.; LACERDA, M.A.S.; SILVA, W.J.; AMORIM, M.J.A.A.L. Ramos do arco aórtico e da aorta torácica em bicho-preguiça (Bradypus variegatus). Arquivos Brasileiros de Medicina Veterinária e Zootecnia, v. 70, p. 1203-1211, 2018. Disponível em: https://doi.org/10.1590/1678-4162-9963
ALMEIDA, L.B.; QUEIROLO, D.; BEISIEGEL, B.M.; OLIVEIRA, T.G. Avaliação do estado de conservação do Gato-mourisco Puma yagouaroundi (É. Geoffroy Saint-Hilaire, 1803) no Brasil. BioBrasil, v. 3, n. 1, p. 99-106, 2013. Disponível em: https://doi.org/10.37002/biodiversidadebrasileira.v3i1.376
BIIHRER, D.A.; GUIMARÃES, G.C.; LOPES, G.C.; LIMA, I.G. Descrição anatômica dos ramos arteriais da aorta torácica e abdominal do quati (Nasua nasua) (Carnivora, Procyonidae). Biotemas, v. 28, n. 2, p. 119-124, 2015. Disponível em: https://doi.org/10.5007/2175-7925.2015v28n2p119
BUDRAS, K.D.; MCCARTHY, P.H.; FRICKE, W.; RICHTER, R.; HOROWITZ, A.; BERG, R. Anatomia do Cão – texto e atlas. 5 ed. São Paulo: Manole, 2012. 219 p.
CAMPOS, R.; DE ARAÚJO, A.C.P.; DE AZAMBUJA, R.C. Ramos colaterais do arco aórtico e suas principais ramificações em nutria (Myocastor coypus). Acta ScientiaeVeterinariae, v. 38, n. 2, p. 139-146, 2010. Disponível em: https://doi.org/10.22456/1679-9216.16612
CASO, A.; OLIVEIRA, T.; CARVAJAL, S.V. Herpailurus yagouaroundi. The IUCN Red List of Threatened Species 2015: e.T9948A50653167. Disponível em: http:// dx.doi.org/10.2305/IUCN.UK.2015-2.RLTS.T9948A50653167.en.
CUBAS, Z.S.; SILVA, J.C.R.; CATÃO-DIAS, J.L. Tratado de Animais Selvagens. 2 ed. São Paulo: Roca, 2014. p. 870- 87.
CULAU, P.O.V.; DE AZAMBUJA, R.C.; CAMPOS, R. Ramos colaterais viscerais da artéria aorta abdominal em Myocastor coypus (nutria). Acta ScientiaeVeterinariae, v. 36, n. 3, p. 241-247, 2008. Disponível em: https://doi.org/10.22456/1679-9216.17293
DE OLIVEIRA, R.E.M.; DE ARAÚJO JÚNIOR, H.N.; DA SILVA COSTA, H.; DE OLIVEIRA, G.B.; DE MOURA, C.E.B.; DE MENEZES, D.J.A.; DE OLIVEIRA, M.F. Artérias colaterais do arco aórtico do gerbil (Meriones unguiculatus) Acta Scientiae Veterinariae, v. 46, p. 1609, 2018. Disponível em: https://doi.org/10.22456/1679-9216.79176.85987
DE SOUZA MARQUES, I.C.; GARCIA FILHO, S.P.; MARTINS, L.L.; LEAL, L.M., DOS REIS, A.C.G.; MACHADO, M.R.F. Artérias mesentéricas cranial e caudal da paca (Cuniculus paca, L. 1766). Biotemas, v. 26, n. 1, p. 165-171, 2013. Disponível em: https://doi.org/10.5007/2175-7925.2013v26n1p165
DYCE, K.M.; SACK, W.O.; WENSING, C.J.G. Tratado de Anatomia Veterinária. 5 ed. Rio de Janeiro: Editora Guanabara Koogan, 2019. 872 p.
FARIA, B.M.; BRANCO, É.; LIMA, A.R. Ramos da aorta abdominal de Aotus azarae infulatus. Biotemas, v. 29, n. 2, p. 69-76, 2016. Disponível em: https://doi.org/10.5007/2175-7925.2016v29n2p69
FELIPE, R.; EURIDES, D.; SILVA, D.C.; SILVA, Z.; CHACUR, E.; BARROS, R.; SILVA, D.R. Segmento caudal da artéria aorta abdominal e suas ramificações em quati (Nasua nasua) Linnaeus, 1766. Enciclopedia Biosfera, v. 4, n. 26, 2017. Disponível em: https://doi.org/10.18677/EnciBio_2017B90
FURTADO, D.F.S.; VASCONCELOS, L.D.P.; BRANCO, É.; LIMA, A.R. Anatomia cardíaca e ramificações da aorta em macaco-prego (Sapajus apella). Biotemas, v. 30, n. 4, p. 83-93, 2017. Disponível em: https://doi.org/10.5007/2175-7925.2017v30n4p83
GERRITY, L.W.; MARTIN, J.E. Anomalous right subclavian artery in a dog. The Southwestern Veterinarian, College Station, v. 33, n. 1, p. 76-77, 1980.
GHOSHAL, N.G. Coração e artérias do carnívoro. In: GETTY R. (Ed.). Anatomia dos animais domésticos. 5 ed. Rio de Janeiro: Interamericana, 1986. p. 1497-1550.
GOMES, A.R.A.; SILVA, F.O.C.; RIBEIRO, L.A.; DE SOUZA, T.A.M. Origens e distribuições das artérias mesentéricas cranial e caudal em fetos de javalis (Sus scrofa scrofa, Linnaeus–1758). Biotemas, v. 26, n. 1, p. 159-164, 2013. Disponível em: https://doi.org/10.5007/2175-7925.2013v26n1p159
KONIG, E.H.; LIEBICH, H. Anatomia dos animais domésticos: texto e atlas colorido. 7. ed. Porto Alegre: Artmed, 2021. 856 p.
LEÃO, L.; SOUZA, E.C.; DE CARVALHO, A.D.; DE SOUZA JUNIOR, P. Topografia da artéria celíaca no Lycalopex gymnocercus (Cuvier, 1798) Anais do Salão Internacional de Ensino, Pesquisa e Extensão, v. 9, n. 2, 2017.
LIMA, A.R.; SOUZA, D.C.D.; CARMO, D.C.D.; SANTOS, J.T.; BRANCO, É. Ramos colaterais do arco aórtico e suas principais ramificações no cachorro-do-mato (Cerdocyon thous). Pesquisa Veterinaria Brasilica, v. 36, p. 647-651, 2016. Disponível em: https://doi.org/10.1590/S0100-736X2016000700015
MACEDO, B.C.; LIMA, A.R.; PEREIRA, L.C.; BRANCO, É. Descrição morfológica dos ramos colaterais da aorta abdominal do tamanduá-mirim (Tamandua tetradactyla). Biotemas, v. 26, n.1, p. 173-180, 2013. Disponível em: https://doi.org/10.5007/2175-7925.2013v26n1p173
MARTINS, D.M.; LIMA, A.R.; PINHEIRO, L.L.; DE SANTA BRÍGIDA, S.S.; ARAÚJO, E.B.; MELUL, R.; BRANCO, É. Descrição morfológica dos ramos colaterais do arco aórtico e suas principais ramificações em Leopardus pardalis. Acta Veterinaria Brasilica, v. 4, n. 2, p. 74-77, 2010. Disponível em: https://periodicos.ufersa.edu.br/index.php/acta/article/view/1736
MICHALSKI, F.; CRAWSHAW, P.G.; OLIVEIRA, T.G.D.; FABIÁN, M.E. Notes on home range and habitat use of three small carnivore species in a disturbed vegetation mosaic of southeastern Brazil/Notes sur le territoire et l'utilisation de l'habitat de trois espèces de petits carnivores dans une végétation mosaïque perturbée au Sud Est du Brésil. Mammalia, v.70, p. 52-57, 2006. Disponível em: https://doi.org/10.1515/MAMM.2006.004
NETO, L.F.L.; SOUZA, E.C.; DOS SANTOS, E.A.R.; MONTANA, M.M.; CARVALHO, A.D.; JUNIOR, P.S. Esqueletopia e topografia da artéria celíaca no Lycalopex gymnocercus (Fischer, 1814). Revista Acadêmica Ciência Animal, v.17, p. 2596-2868, 2019. Disponível em: https://doi.org/10.7213/1981-4178.2019.17007
OLIVEIRA, R.E.; OLIVEIRA, G.B.; BARBOSA, P.M.; BEZERRA, F.V.; ALBUQUERQUE, J.F.G.D.; AMBRÓSIO, C.E.; OLIVEIRA, M.F. Ramos colaterais do Arco aórtico do preá (Galea spixii Wagler, 1831). Pesquisa Veterinária Brasileira, v. 35, p. 762-766, 2015. Disponível em: https://doi.org/10.1590/S0100-736X2015000800008
PEREIRA, K.F.; DE SOUZA TERRA, D.R.; FERREIRA, L.S.; SABEC-PEREIRA, D.K.; LIMA, F.C.; DOS SANTOS, O.P. Descrições anatômicas do coração e vasos da base de Procyon cancrivorus (CUVIER, 1798). Arquivos do MUDI, v. 20, n.3, p. 1-12, 2016. Disponível em: https://doi.org/10.4025/arqmudi.v20i3.34343
PINHEIRO, L.L.; ARAÚJO, E.B.; LIMA, A.R.; MARTINS, D.M.; MELUL, R.; SOUZA, A.C.B.; BRANCO, É. Os ramos colaterais da aorta abdominal em jaguatirica (Leopardus pardalis). Pesquisa Veterinária Brasileira, v.34, p. 491-495, 2014. Disponível em: https://doi.org/10.1590/S0100-736X2014000500018
PINHEIRO, V.D.L.C.; DE LIMA, A.R.; PEREIRA, L.C.; GOMES, B.D.; BRANCO, É. Descrição anatômica dos ramos colaterais do arco aórtico do tamanduá-mirim (Tamandua tetradactyla). Biotemas, v. 25, n. 2, p. 133-137, 2012. Disponível em: https://doi.org/10.5007/2175-7925.2012v25n2p133
QUEIROZ, P.V.; MOURA, C.E.; LUCENA, J.Á.; OLIVEIRA, M.F.; ALBUQUERQUE, J.F. Artérias mesentéricas cranial e caudal em mocós Kerodon rupestris (Wied, 1820). Pesquisa Veterinária Brasileira, v. 31, p. 623-626, 2011. Disponível em: https://doi.org/10.1590/S0100-736X2011000700013
ROCHA, E.F.; DOS SANTOS, N.T.A.; DIAS, R.F.F.; DINIZ, J.A.R.A.; DOS SANTOS, J.R.S.; DE MENEZES, D.J.A. Anatomia macroscópica dos órgãos reprodutores do Puma yagouaroundi (Geoffroy, 1803) macho. Pubvet, v. 11, p. 744-839, 2017. Disponível em: https://doi.org/10.22256/pubvet.v11n8.767-770
SANTOS, A.L.Q.; MUNDIM, A.V.; MORAES, F.M.; ALVARENGA, G.J.R.; MARQUES, F.K.; BORGES, K.M.; JÚNIOR, J.Á. Origem e ramificações das artérias mesentéricas cranial e caudal em tartaruga da Amazônia Podocnemis expansa Schweigger, 1812, (Testudinata-Pelomedusidae). Archives of Veterinary Sciences, v. 9, n. 2, p. 49-53, 2004. Disponível em: http://dx.doi.org/10.5380/avs.v9i2.4064
SILVA, B.X.; ROZA, M.S.; HERNANDEZ, J.M.F.; DE SOUZA, H.J.M.; FIGUEIREDO, M.A. Artéria lienal em gatos: estudo aplicado à pesquisa anatômica e a prática cirúrgica. Brazilian Journal of Veterinary Medicine, v. 33, n.1, p. 41-47, 2011. Disponível em: https://www.researchgate.net/publication/286327986_Lienal_artery_in_cat_Applied_study_for_anatomical_research_and_surgical_practice
TÓFOLI, C.F.; ROHE, F.; SETZ, E.Z.F. Jaguarundi (Puma yagouaroundi) (Geoffroy, 1803) (Carnivora, Felidae) food habits in a mosaic of Atlantic Rainforest and eucalypt plantations of southeastern Brazil. Brazilian Journal of Veterinary Research and Animal Science, v.69, p. 871-877, 2009. Disponível em: https://doi.org/10.1590/S1519-69842009000400015
VITUMS, A. Anomalous origin of the right subclavian and common carotid arteries in the dog. Cornell Veterinarian, v.52, p. 5-15, 1962.
XAVIER-SILVA, B.; ROZA, M.S.; BABINSKI, M.A.; PALHANO, H.B.; ABIDU-FIGUEIREDO, M. Ramos principais da artéria celíaca no gato doméstico: estudo aplicado à pesquisa experimental e à prática cirúrgica. Revista Brasileira de Ciências Veterinárias, v.19, n. 3, p. 133-138, 2012. Disponível em: https://doi.org/10.4322/rbcv.2014.104
XAVIER-SILVA, B.; ROZA, M.S.; BABINSKI, M.A.; SCHERER, P.O.; PALHANO, H.B.; ABIDU-FIGUEIREDO, M. Morfometria, origem e esqueletopia da artéria celíaca no gato doméstico. Brazilian Journal of Veterinary Medicine, v. 35, n. 3, p. 253-259, 2013. Disponível em: https://bjvm.org.br/BJVM/article/view/617.
Recebido em: 18/09/2024
Publicado em: 08/05/2025